菁于教,优于学
旗下产品
校本题库
菁优备课
开放平台
菁优测评
菁优公式
天天优学
菁优App
数字备考
菁优阅卷
充值服务
试卷征集
申请校本题库
智能组卷
免费试用
错题库
免费试用
五大核心功能
组卷功能
资源共享
在线作业
在线测评
试卷加工
登录
进入游客模式
为了更流畅的使用菁优平台,请将您浏览器的极速模式切换为兼容模式,或使用IE8及以上版本的浏览器
关闭
温馨提示:如果您的页面出现按钮不可点或局部遮挡现象,请将浏览器切换到极速模式或ie浏览器升级到“ie8”以上!
试题
试题
试卷
搜单题
搜整页
请选取单道试题图片,将图片拖拽或粘贴到这里
上传图片文件
(图片大小不超过5M)
搜题
清空
试卷征集
加入会员
操作视频
试题搜索
同城
热搜榜
|
组卷
有奖答题
当前位置:
试题搜索
>
同城
科目:
小学数学
小学语文
小学英语
小学奥数
小学科学
小学道德与法治
初中数学
初中物理
初中化学
初中生物
初中科学
初中地理
初中语文
初中英语
初中道德与法治
初中历史
高中数学
高中物理
高中化学
高中生物
高中地理
高中语文
高中英语
高中政治
高中历史
高中信息
高中通用
初中信息技术
中职数学
中职语文
中职英语
题型
全部题型
选择题
填空题
计算题
解答题
题类
全部
常考题
易错题
好题
压轴题
难度
全部
易
较易
中档
较难
难
年份
全部
2026
2025
2024
2023
2022
来源
全部
杯赛真题
杯赛模拟
地区竞赛
学校竞赛
竞赛专题
竞赛训练
共
21
条信息
一位妇女W、她的兄弟B、儿子
S
和女儿D(所有的关系都是血缘关系,兄弟B和儿子
S
和女儿D年龄可以相等)都是棋手。最坏的棋手的孪生者(也是四个棋手之一)和最好的棋手为异性,最坏的棋手和最好的棋手为同年龄的。则最坏的棋手是( )
A.女儿D
B.妇女W
C.儿子
S
D.兄弟B
E.其它都不对
更新:2025/12/18
组卷:0
引用:1
难度:0.5
解析
纠错
收藏
相似题
下载
提示:下载将消耗您1次下载权益,建议加入试题篮统一下载
试题篮
下表是同一个月的日历的一部分。如果其中一个字母表示的日期与C表示的日期相加恰好等于A和B所表示的日期的和,那么,这个字母是( )
星期二
星期三
星期四
星期五
星期六
C
A
Q
S
B
P
T
R
A.P
B.
S
C.Q
D.T
E.R
更新:2025/12/11
组卷:1
引用:1
难度:0.5
解析
纠错
收藏
相似题
下载
提示:下载将消耗您1次下载权益,建议加入试题篮统一下载
试题篮
(2024•其他杯赛)
如图,沿着圆周依次标出A,B,P,C,Q,D,R,E,
S
,F,使得ABCDEF是一个等边六边形,APQRS 是一个等边五边形,则∠BAP的度数为( )
A.3°
B.5°
C.6°
D.10°
E.15°
《2024年第五届广东省深圳市”超常思维“六年级数学竞赛试卷》
更新:2025/4/28
组卷:23
引用:1
难度:0.5
解析
纠错
收藏
相似题
下载
提示:下载将消耗您1次下载权益,建议加入试题篮统一下载
试题篮
(2024•其他杯赛)
如图,共有6个等边三角形和19个圆圈。如果将1-19这19个自然数分别填入图中的圆圈中,使得每个等边三角形的每条边上的3个数之和都等于
S
,那么
S
的最小值是( )
A.22
B.25
C.27
D.29
E.30
《2024年第五届广东省深圳市”超常思维“六年级数学竞赛试卷》
更新:2025/4/28
组卷:13
引用:1
难度:0.5
解析
纠错
收藏
相似题
下载
提示:下载将消耗您1次下载权益,建议加入试题篮统一下载
试题篮
(2022•其他杯赛)
设
S
=145678+456781+567814+678145+781456+814567,则
S
是( )个不同质数之积。
A.6
B.5
C.4
D.3
E.以上都不对
更新:2025/4/12
组卷:8
引用:1
难度:0.5
解析
纠错
收藏
相似题
下载
提示:下载将消耗您1次下载权益,建议加入试题篮统一下载
试题篮
(2023•其他杯赛)
如图是一种电脑射击游戏的示意图:线段CD、EF和G的长度都是20cm,O、P、Q分别是它们的中点,并且位于同一条线段AB上,AO=45cm,OP=PQ=20m。已知CD上小圆环的速度是5cm/
s
,EF上小圆环的速度是9cm/
s
,GH上小圆环的速度是27cm/
s
。零时刻,CD、EF、GH上各有一个小圆环从左端点同时开始在线段上匀速往返运动,此时,从点A向B发射一颗匀速运动的子弹,要想穿过三个圆环,子弹的速度最大为( )cm/
s
。
A.2.5
B.3
C.3.2
D.4.5
E.5.1
《2023年广东省深圳市“超常思维”六年级数学竞赛试卷》
更新:2024/6/18
组卷:18
引用:1
难度:0.5
解析
纠错
收藏
相似题
下载
提示:下载将消耗您1次下载权益,建议加入试题篮统一下载
试题篮
(2023•其他杯赛)
一个电子装置每60
s
发出“哗”的一声,另一电子装置每62
s
发出“哗”的一声,两者在上午10:00同时发出“哗”声,下一次一起发出“哗”声时的时间是上午( )
A.10:30
B.10:31
C.10:59
D.11:00
E.11:03
《2023年广东省深圳市“超常思维”六年级数学竞赛试卷》
更新:2024/6/18
组卷:43
引用:1
难度:0.5
解析
纠错
收藏
相似题
下载
提示:下载将消耗您1次下载权益,建议加入试题篮统一下载
试题篮
能正确表示图(a),(b),(c),中阴影部分面积
S
A
,
S
B
,
S
C
的大小关系的是( )(图中长度单位为cm)
A.
S
A
>
S
B
>
S
C
B.
S
C
>
S
A
>
S
B
C.
S
A
>
S
C
>
S
B
D.
S
A
=
S
C
>
S
B
E.
S
C
>
S
B
>
S
A
《2022年广东省深圳市“超常思维”五年级数学竞赛试卷》
更新:2024/3/19
组卷:13
引用:1
难度:0.5
解析
纠错
收藏
相似题
下载
提示:下载将消耗您1次下载权益,建议加入试题篮统一下载
试题篮
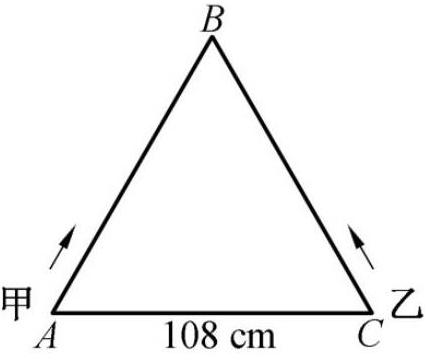
如图,△ABC是边长为108cm的等边三角形,虫子甲和乙分别从点A和点C同时出发,沿△ABC的边爬行。甲顺时针爬行,乙逆时针爬行,速度比是4:5。相遇后,甲在相遇点休息10
s
,然后继续以原来的速度沿原方向爬行;乙不休息,速度提高20
%
,仍沿原方向爬行,第二次恰好在BC的中点相遇.则开始时,虫子甲的爬行速度为( )cm/
s
。
A.8.6
B.9.6
C.10.6
D.11.6
E.15.9
《2022年广东省深圳市“超常思维”五年级数学竞赛试卷》
更新:2024/3/19
组卷:25
引用:1
难度:0.5
解析
纠错
收藏
相似题
下载
提示:下载将消耗您1次下载权益,建议加入试题篮统一下载
试题篮
如图,这是一张5行5列的方格表,顶上一行填有符号P,Q,R,
S
,T,第四行中间填有符号P,Q,R,余下的方格中可填入P,Q,R,
S
,T,要求做到同一符号在每一行、每一列、及每条对角线上只出现一次.那么填入画有阴影的方格中的符号必须是( )
A.P
B.Q
C.R
D.
S
E.T
《2022年广东省深圳市“超常思维”五年级数学竞赛试卷》
更新:2024/3/19
组卷:11
引用:1
难度:0.5
解析
纠错
收藏
相似题
下载
提示:下载将消耗您1次下载权益,建议加入试题篮统一下载
试题篮
1
2
3
下一页
尾页
试卷热搜榜
更多
1
2025年华罗庚金杯六年级思维挑战奥数模拟试卷
2
2026年江苏省我爱学数学杯未来数学家创新人才培养活动二年级数学试卷(1月份)
3
2026年迎春杯数学花园探秘科普活动决赛试卷(三、四年级)
4
2025-2026学年广东省汕尾市六年级(上)竞赛数学试卷
5
2025年第33届WMO世界奥林匹克数学竞赛初赛奥数试卷(六年级)
6
2025年江苏省南京市秋季资优生数学沃土计划活动创新能力测评六年级奥数试卷
7
2026年江苏省我爱学数学杯未来数学家创新人才培养活动六年级数学试卷(1月份)
8
2026年江苏省我爱学数学杯未来数学家创新人才培养活动四年级数学试卷(1月份)
9
2026年世界少年奥林匹克思维能力测评活动(地区赛)五年级数学试卷
10
2026年江苏省我爱学数学杯未来数学家创新人才培养活动一年级数学试卷(1月份)
0
试题篮
题量
0
/0
难度
题量
占比
下一步
我知道了
总计
0
题
清空全部
进入组卷中心
暂无任何试题
试
题
篮
0/0
【新增试题篮】
×
试题篮名称:
设为默认试题篮
确认
取消
试卷征集
商务合作
服务条款
走进菁优
课件征集
帮助中心
兼职招聘
意见反馈
深圳市菁优智慧教育股份有限公司
粤ICP备10006842号
公网安备44030502001846号
©2010-2026 jyeoo.com 版权所有
深圳市市场监管
主体身份认证
APP开发者:深圳市菁优智慧教育股份有限公司
|
应用名称:菁优网
|
应用版本:5.2.7
|
隐私协议
|
第三方SDK
|
用户服务条款
广播电视节目制作经营许可证
|
出版物经营许可证
|
网站地图
本网部分资源来源于会员上传,除本网组织的资源外,版权归原作者所有,如有侵犯版权,请立刻和本网联系并提供证据,本网将在三个工作日内改正





{kind=link}
{kind=link}